Superstruktur der DNA
Die menschliche DNA einer diploiden Zelle besteht aus ungefähr 2 x 3 x 109 Basenpaaren, verteilt auf 46 Chromosomen. Jeder Zellkern enthält somit 46 DNA-Moleküle unterschiedlicher Grösse (vgl. DNA in Zellorganellen wie Mitochondrien und Chloroplasten). Würde man die DNA in den 46 Chromosomen zu einer einzigen Doppelhelix zusammenfügen, käme man auf eine Länge von 2 Metern DNA pro Zelle! Es ist einleuchtend, dass die DNA Strukturen annehmen muss, die es erlauben, sie in den Zellkern zu packen und sie vor Scherkräften zu schützen, die die nur 2 nm Dicke Doppelhelix in Stücke reissen könnten. Aus diesem Grund binden bestimmte Proteine an die DNA und verformen diese zu stabilen Superstrukturen. Solche DNA-Proteinkomplexe nennt man Chromatin. Bei Zellteilungen - Mitosen und Meiosen -, müssen sich die Chromosomen zu Mitosechromosomen maximal verdichten (kondensieren), damit sie überhaupt auf die Tochterzellen verteilt werden können. Betrachten wir also vorerst die sogenannten Interphasenchromosomen.
Die an DNA gebundenen Proteine machen einen beträchtlichen Teil des Chromatins aus. Tatsächlich besteht Chromatin etwa zur Hälfte aus Proteinen, welche noch in Histone und Nicht-Histon-Proteine unterteilt werden können.
Histone sind kleine, basische Eiweisse, die nur in Eukaryonten vorkommen. Mit ihrem hohen Anteil an positiv geladenen Aminosäuren wie Arginin und Lysin können sie relativ unspezifisch an die DNA binden und die negativen Ladungen der Phosphatgruppen des "DNA-Rückgrats" kompensieren.
Überlegen Sie sich, welche Auswirkungen es auf die Stabilität der DNA hätte, wenn die negativen Ladungen der Helix nicht aufgehoben würden.
Lösung
X
Die negativen Ladungen würden sich gegenseitig abstossen und die Doppelhelix destabilisieren.
Es gibt insgesamt fünf verschiedene Klassen von Histonen: H1, H2A, H2B, H3 und H4. Gewisse eukaryontische Zellen - wie z.B. die Erythrocyten des Huhns - haben noch einen weiteren Typ, das Histon H5, welches als Variante von H1 angesehen wird. Je zwei Moleküle H2A, H2B, H3 und H4 verbinden sich zu einem Histon-Octamer. Die DNA-Doppelhelix wickelt sich nun in Linkswindungen um dieses Octamer wie ein Faden um eine Spule. Die dadurch entstehende Struktur ist das Nucleosom.
Interessierte finden hier
X
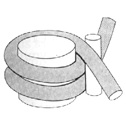
Core-Partikel: Die Doppelhelix wickelt sich 1 3/4-mal um das Histon-Oktamer. Dies entspricht 146 Basenpaaren (bp) DNA.
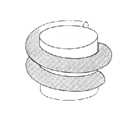
Chromatosom: Es enthält zwei volle Windungen DNA (166 bp DNA). Daran bindet zusätzlich noch das Histon H1 (vertikaler Zylinder, rechts).
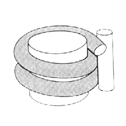
Nukleosom: Es besteht ebenfalls aus zwei Windungen DNA, dem Histon-Octamer und H1, hat aber noch sog. Linker-DNA von etwa 30 bp, welche benachbarte Nukleosomen miteinander verbindet. Ein Nukleosom enthält also etwa 200 bp DNA.
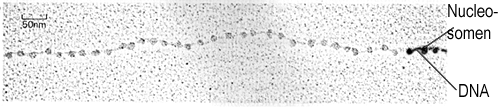
Behandelt man Chromatin mit einer stark hypotonen Lösung, so lassen sich die Chromosomen als Nukleosomenfäden im Elektronenmikroskop darstellen. Sie gleichen einer Perlenkette:
Archiv AUM
1977
Die nächsthöhere strukturelle Einheit des Chromatins ist das Solenoid. Für die Ausbildung des Solenoids ist das Histon H1 essentiell: es verbindet benachbarte Nukleosomen zu einer 30 nm dicken Chromatinspirale. Wie sich dieser Prozess genau vollzieht, ist aber noch weitgehend unklar. Folgendes EM-Bild veranschaulicht die dichte Packung der Nukleosomen:
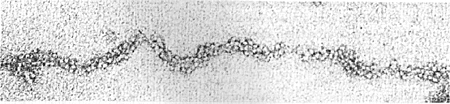
Archiv AUM
1977
Die Solenoid-Faser wird gelegentlich von Nicht-Histon-Proteinen in Segmente unterteilt, wie folgendes Modell zeigt:
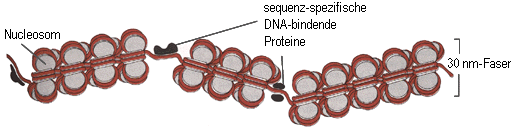
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle", VCH, 2. Auflage 1990,
ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
Unten ein Schema, das das Prinzip der Faltungen wiedergibt:
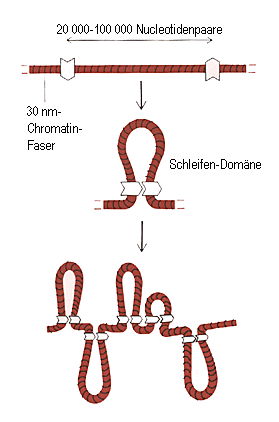
Bildquelle: Alberts et al,: "Molekularbiologie der Zelle",
VCH, 2. Auflage 1990, ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
Hinweis: D. Hewish und L. Burgoyne waren unter den ersten, welche sich mit der Strukturaufklärung des Chromatins beschäftigten.
Sowohl in Eukaryonten wie in Prokaryonten gibt es Enzyme, die Topoisomerasen, die unter ATP-Verbrauch einen oder beide DNA-Stränge auftrennen, durch Rotation um die Helixachse die Windungszahl pro Längeneinheit ändern und dann die freien Enden wieder verknüpfen (ligieren). Die Topoisomerase I spaltet dabei nur einen, die Topoisomerase II beide DNA-Stränge. Nimmt die Windungszahl pro Längeneinheit zu, spricht man von positivem Supercoiling, nimmt sie ab, von negativem Supercoiling. Diese Konformationsänderung setzt die DNA unter Torsionsspannung. Hat die DNA ein freies Ende, so kann sie sich wieder entspannen, indem sie um ihre Achse rotiert. Sind dagegen beide Enden fixiert, wie bei der Ring-DNA der Prokaryonten oder bei Chromatin-Schleifen, kann die DNA die Überspannung nur durch Ausbildung einer superhelicalen Schleife kompensieren (zwei fixe Enden können nicht frei rotieren). Bei dieser DNA-Superhelix ist die DNA zusätzlich zu den Doppelhelixwindungen umeinander verdrillt:
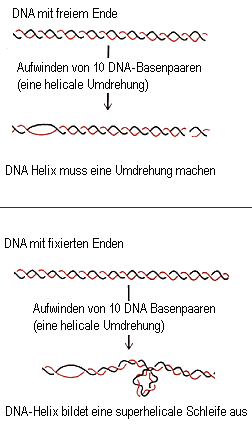
Bildquelle: Alberts et al,:
"Molekularbiologie der Zelle",
VCH, 2. Auflage 1990,
ISBN 3-527-27983-0
und 3. Auflage 1995, ISBN 3-527-30055-4
Sie können sich die Entstehung dieser Konformation am besten vorstellen, wenn sie an ein spiraliges Telefonkabel denken, welches Sie aus einem gedehnten in einen entspannten Zustand führen.
Zu Replikations-Elementen: DNA-Topoisomerasen
Hinweis: Gute Dienste leistet ein Schnur-Modell
X
Anleitung zum Schnurmodell
1. Spannen Sie einen ca. 10 cm grossen Abschnitt der Schnur zwischen Daumen und Zeigefinger Ihrer beiden Hände:

2. Verdrillen Sie ein Ende mit Daumen und Zeigefinger der geschickteren Hand, während Sie das andere Ende festhalten. Die Windungszahl der Schnur hat nun entweder zugenommen (positives Superscrolling) oder abgenommen (negatives Superscrolling):
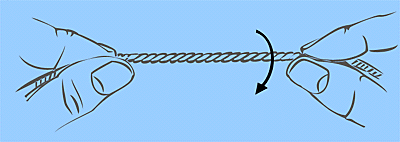
3. Entlasten Sie nun die Spannung auf der Schnur, indem Sie die beiden Enden einander nähern. Die Schnur wird dadurch superhelicale Windungen erfahren:
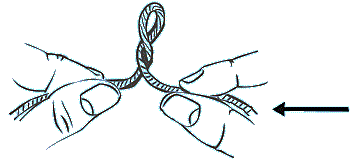
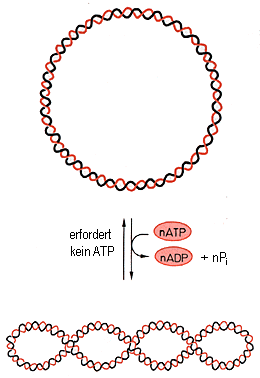
Topoisomerasen benötigen für die Rückführung (Relaxation) in den Normalzustand kein ATP; die Energie steckt nämlich in der Supercoil-Konformation selbst:
Karlson, Doenecke, Koolman:
"Kurzes Lehrbuch der Biochemie für Mediziner und Naturwissenschaftler",
Georg Thieme Verlag Stuutgart, 14. neubearbeitete Auflage 1994,
ISBN 3-13-357814-6
Negatives Supercoiling erleichtert wahrscheinlich Prozesse wie Replikation oder Transkription, weil die "unterwundene" DNA den Zugang bestimmter Proteine zu Steuersequenzen erleichtert.